
Saturação das bases
A saturação das bases é um dos principais parâmetros utilizados em análises de solos. É a relação entre a quantidade dos cátions básicos em relação aos cátions ácidos que compõe a CTC a ph 7.
Bases: são cátions, que em sua hidrólise liberam íons hidroxilas.
Ácidos: são cátions, que na hidrolise liberam H3O+ e o próprio íon hidrogênio (H+)
Nos solos, grosso modo, se tem duas fontes de íons que regulam sua acidez.
Fontes do Íon Hidrogênio
O complexo coloidal do solo (minerais de argila e matéria orgânica) pode ser considerado como uma mistura de compostos insolúveis como as próprias micelas e solúveis como ácidos e seus sais.
O hidrogênio e outros cátions metálicos ao redor das micelas insolúveis permanecem em equilíbrio dinâmico com a solução do solo. Portanto, o ph é altamente influenciado por estes cátions adsorvidos à medida que ocorre a dissociação.
Há duas fontes de acidez ou de íons de hidrogênio para a solução do solo;
- Íons de hidrogênio propriamente ditos.
- Íons de hidrogênio provenientes da hidrólise dos monômeros do alumínio.
Em condições muito ácidas o alumínio adsorvido se torna solúvel e fica presente em solução na forma iônica como Al3+. Como são trivalentes ficam adsorvidos às micelas com maior avidez, assim como os íons de hidrogênio. Dessa forma, um solo muito ácido é dominado por Al3+ e H+.
O alumínio adsorvido está em equilíbrio com o Al+3 da solução.
A reação mostra como o alumínio adsorvido pode aumentar a acidez do solo.
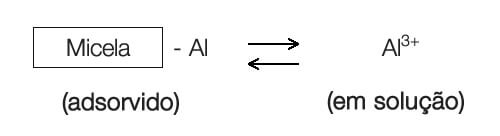
Uma vez em solução o Al3+ pode ser hidrolisado da seguinte maneira:
Al3+ + 3H2O → Al (OH)3 + 3H+
O Al(OH)3 é insolúvel, portanto não hidrolisável, sobrando, no entanto, o H+ que é liberado na solução provocando o abaixamento do pH. Esse é o principal motivo pelo qual o alumínio, em última análise acidifica o solo.
Este tipo de acidez é conhecido por acidez trocável, uma vez que, o alumínio era trocável na CTC.
Assim, o efeito do alumínio e do próprio hidrogênio adsorvidos é aumentar a concentração de íons Hidrogênio (H1) na solução, consequentemente reduzindo o pH da mesma.
Origem dos Íons Hidroxílicos (OH–)
Se o hidrogênio e o alumínio adsorvidos forem substituídos por outros cátions como CÁLCIO, MAGNÉSIO, POTÁSSIO e SÓDIO, diminuirão a concentração de íons H+ na solução do solo.
A presença destes cátions aumentará a concentração de íons OH– em solução, uma vez que, há uma relação inversa entre H+ e OH–.
Assim os cátions são conhecidos como formadores de bases (OH–) se transformam em fontes de OH– apenas pela substituição do Hidrogênio adsorvido.
Portanto os cátions metálicos, Ca, Mg, K e Na exercem influência direta na concentração de íons OH– na solução do solo.
A reação desses cátions adsorvidos aos coloides resulta em reação completamente alcalina.
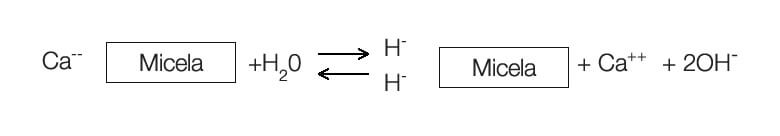
Por isso estes cátions (Ca, Mg, K e Na) são chamados de bases. Na verdade a sua reação é alcalina, portanto formadores de bases.
Assim, o pH da solução do solo dependerá da quantidade de H+ e OH– portanto, das quantidades relativas de íons hidrogênio adsorvidos e da quantidade dos cátions metálicos adsorvidos.
Porcentagem da saturação das bases
Verifica-se que para aumentar a saturação das bases para contrapor-se à presença de H+ e Al+++ deve se adicionar cátion metálicos como cálcio, magnésio e potássio.
Este feito leva a um contraponto do pH do sistema. Ressalta-se que o pH não é fator apenas da presença das bases, mas há outros fatores de influência como, por exemplo: a natureza da micela, isto é, se são oxídica, silicatada, húmica e outras.
Dos principais cátions solúveis que se encontram nos solos os mais comuns são fornecidos pela calagem, método usado para fornecer ânions neutralizantes como o bicarbonato (HCO3–) e a hidroxila (OH–), e os cátions cálcio e/ou magnésio.
Dessa forma, a adição de cátions metálicos básicos, é a chave para regulação do pH. Por outro lado, esses cátions metálicos mais comuns como Ca+, Mg+ e K+ são considerados nutrientes essenciais.
Assim, abaixo se relata como estes cátions influem na nutrição das plantas.
Para que servem as bases para as plantas?
Cálcio
O cálcio é um cátion bivalente com raio hidratado de 0,412nm, com alta energia de hidratação. Sua concentração nas plantas pode atingir até 10% do conteúdo de matéria seca dos tecidos, principalmente em folhas mais velhas e tecidos lenhosos sem provocar fitotoxicação ou inibição do crescimento.
Abaixo estão relacionados fatores fisiológicos afetados pelo cálcio.
1. O cálcio é o elemento fundamental do apoplasto (paredes celulares), está sempre ligado as cadeias de ácidos poli-galacturônicos (conjunto conhecido como pectinas) formando complexos bastante estáveis de pectato de cálcio.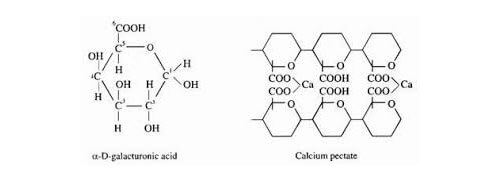
2. O cálcio é o principal elemento de ligação dos grupos de fosfolipídios das membranas celulares. É o elemento que da estabilidade a estas ligações, portanto responsável pela estabilidade das membranas e consequentemente sobre sua seletividade. Sua falta causa a desintegração das membranas e perda da compartimentalização celular. A salinidade induzida por (Na+ e K+), compete com o cálcio neste papel levando a desintegração da membrana plasmática e ao estresse salino.
3. Na maioria das espécies as maiores concentrações de cálcio se encontram na lamela média das paredes celulares e nas superfícies exteriores das membranas celulares fazendo a ponte com os fosfolipídios dando estabilidade tanto as paredes quanto as membranas. Portanto, é o principal elemento na Capacidade de Troca de Cátions (CTC) nas paredes celulares das raízes.
4. Ao contrário do vacúolo a concentração de cálcio é mantida no citoplasma em níveis muito baixos. Estas concentrações baixas no citoplasma têm relação com a disponibilidade de fósforo, pois senão haveria uma provável precipitação de fosfato como fosfato de cálcio insolúvel, diminuindo sua disponibilidade e levando a deficiência de fósforo. Assim o caminho preferencial do cálcio é o acúmulo no vacúolo.
5. O principal transportador de cálcio nas membranas é uma bomba de cálcio tipo antiporte (Ca++ / H+) ATPase, portanto com consumo de ATP.
6. Grande parte cálcio nos cloroplastos se encontram associados às membranas dos tilacóides, com baixa concentração no estroma.
7. A enzima (frutose,1,6 bisfosfato) que regula a síntese de sacarose no citoplasma é afetada pelo cálcio. Concentrações de cálcio livre menores que 1 μM (micro Mol) inibem a atividade de enzima, mesmo em altas concentrações de magnésio. Portanto o excesso de cálcio livre compromete a síntese de sacarose que é o principal carbohidrato solúvel e livre nas plantas.
8. A degradação de pectatos (principal componente de paredes celulares) é mediada pela enzima polygalacturonase que é drasticamente inibida na presença de cálcio, mantendo a integridade das paredes celulares. Por outro lado, em plantas deficientes em cálcio a atividade desta enzima é aumentada, assim o sintoma típico da deficiência de cálcio é a desintegração da parede celular e consequente colapso dos tecidos.
Em plantas adequadamente nutridas em Ca, há grande resistência a infecções por bactérias e principalmente fungos.
9. Em frutos durante a maturação o teor de cálcio cai muito, levando ao aumento da atividade da enzima polygalacturonase, assim promovendo a degradação das paredes celulares permitindo que os frutos entrem em maturação. A aplicação de cálcio nesta fase pode retardar a maturação dos frutos.
10. O cálcio é indispensável no solo para o crescimento da ponta da raiz (coifa). O cálcio deve estar livre na solução do solo. Caso haja deficiência o crescimento da raiz cessa. O cálcio é fundamental para manter a mucilagem na coifa da raiz, além de governar o geotropismo. Da mesma forma, para o tubo polínico, o movimento quimiotrópico é governado pelo gradiente de cálcio extracelular.
11. O cálcio juntamente com a giberilina estimulam a enzima α amilase na geminação de sementes de cereais. Além de reduzir o movimento da enzima para as camadas de aleurona, o cálcio é constituinte da enzima α amilase produzida no reticulo endoplasmático. O transporte de Ca++ no reticulo endoplasmático é ativado por giberilinas e inibido por ácido abscísico (ABA).
12. O cálcio na solução externa regula a seletividade na absorção de íons e previne a saída de solutos do citoplasma.
13. A proteção da membrana produzida pela presença do cálcio livre é mais proeminente em condições de baixa temperatura ou congelamento além de condições de anaerobiose. Assim o cálcio alivia os efeitos de baixa temperatura e anaerobiose pela redução nas perdas de solutos das células e do efluxo de potássio. O efeito na diminuição da taxa respiratória sob estresse de temperatura ou oxigênio melhora a taxa de síntese de proteínas.
14. A senescência dos tecidos se deve ao aumento da concentração de radicais livres levando à peroxidação dos lipídeos das membranas. O cálcio protege os tecidos deste efeito deletério juntamente com as citocininas diminuindo a peroxidação dos lipídeos das membranas com a redução da atividade da enzima lipoxygenase.
15. O cálcio não somente protege as membranas, mas também esta envolvida na sua destruição controlada principalmente durante o crescimento de cotilédones durante a geminação como as fosfolipases.
16. O cálcio estimula muitas enzimas ligadas às membranas como as ATPases em raízes responsáveis pela absorção iônica de outros nutrientes.
17. O cálcio é responsável pela regulação dos canais iônicos e pelo bloqueio da bomba de prótons levando a regulação de absorção de potássio como mantenedor do poder de osmorregulação, principalmente nos movimentos de abertura e fechamento de estômatos além de movimentos násticos.
18. No citoplasma o principal alvo dos sinais de cálcio são proteínas chamadas de “proteínas de modulação por cálcio” sendo os mais conhecidos como “calmodulinas” (Ca – M) que podem ser classificadas em cálcio dependentes Kinase e cálcio independentes Kinase. As cálcio dependentes são prontamente ativados por cálcio e essas enzimas promovem a fosforilação de outras enzimas localizadas nas membranas plasmáticas. As principais enzimas estimuladas são:
- Bombas de prótons ATPases,
- NAD Kinase que cataliza a conversão de NAD em NADP com aceptor final de elétrons nos cloroplastos durante a fotossíntese.
19. As dicotiledôneas são mais exigentes em cálcio do que as monocotiledôneas, por exemplo, o tomate exige 40 vezes mais cálcio que o trigo.
Magnésio
O magnésio é um íon divalente com raio iônico de 0,428 mm com alta energia de hidratação.
A sua taxa de absorção pelas plantas pode diminuir muito na presença de potássio, cálcio e arsênio ou por hidrogênio em condições de pH baixo.
As suas principais funções estão descritas abaixo.
1. Sua principal função esta relacionado com sua capacidade de fazer ligações estáveis com ligantes através de ligações iônicas e tornando-se elemento ponte em ligações estáveis como na molécula de “clorofila”, por exemplo, que aliás é uma ligação covalente forte
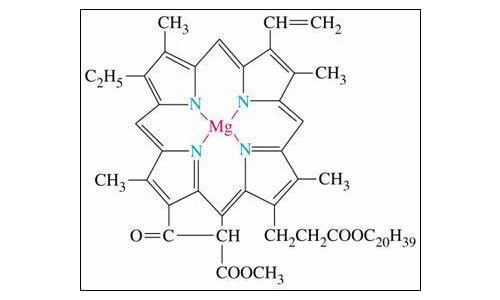
2. O magnésio ainda se posiciona em ligações estáveis de proteínas com pontes estabelecendo ligações geométricas entre as enzimas e o substrato. A Ribulose 1,5, bisfosfato Carboxilase é um exemplo desta ligação.
3. Grande parte do magnésio nas plantas se encontra envolvidos na regulação do pH citoplasmático e no balanço cátions ânions. É também relevante sua participação nas ligações de pectato na parede celular, assim como o caso do cálcio formando pectatos de magnésio.
4. Cerca de 85 % do magnésio nas plantas se encontra na forma livre principalmente nos vacúolos e são usados na regulação osmótica e no balanço cátion – ânion no citoplasma, o restante cerca de 6% a 25% de todo o magnésio na planta se encontram associados as moléculas de clorofila. Cerca de 5 a 10% se encontram na forma de pectatos na parede celular ou precipitado nos vacúolos
5. Clorofila e síntese de proteínas: Na síntese de clorofila o Mg e grupos heme (Fe) tem uma rota de síntese compartilhada até o nível de proto clorofila a partir do precursor, que é o ácido glutâmico. A inserção do magnésio na molécula de porfirina é o primeiro passo para a formação da clorofila e este passo é catalizado pela enzima Mg-Chelatase. Esta enzima requer ATP e assim magnésio adicional.
6. O magnésio tem uma função essencial como elemento de ligação para a agregação de subunidades de ribossomos para a síntese de proteínas. Quando o nível de magnésio livre (Mg++) esta deficiente, ou na presença de excesso de potássio (K+) as sub unidades de ribossomos se dissociam e a síntese de proteínas cessa.
7. O magnésio também é requerido para a RNA-polimerase para a formação de RNA-mensageiro no núcleo da célula.
8. Atuação Enzimática, Fosforilação e Fotossíntese
- Glutationa Sintase.
- PEP-carboxilose – Esta enzima é ativada na presença de .magnésio melhorando a assimilação de CO2 na fotossíntese .das plantas de metabolismo C4
- Fosfatases, ATPases e Caboxilases – Nestas enzimas o Mg é preferencialmente ligado aos grupos de nitrogênio ou fosforil como no caso do ATP. O substrato das ATPases assim como as P Pases é a ligação Mg-ATP que predomina em relação ao ATP sozinho. O complexo Mg ATP é que ativa as enzimas de ATPases como por exemplo a H+ ATPase nas membranas para o fluxo de hidrogênios nas membranas no caso da absorção de íons catiônicos.A Atividade máxima dessas enzimas requerem a presença de Mg++ e K+. Assim a presença de Mg+ livre ativa as enzimas de fosforilação.A própria síntese do ATPFosforilação = ADP + P → ATP tem alto requerimento de Mg++ como elo de ligação entre ADP e a enzima.Por outro lado, a adição de Ca++ inibe severamente a fosforilação.É por isso que a concentração de cálcio Ca++ é mantida muito baixo dentro dos cloroplastos e mitocôndrias onde ocorrem a maior parte das enzimas de fosforilação.
- Ribulose 1.5 bisfosfato carboxilase e oxidase. (RUBISCO)A atividade desta enzima responsável pela assimilação de CO2 no ciclo de Calvin é altamente modulado por Mg++ e pH. A ligação do Mg++ na enzima aumenta a sua afinidade pelo CO2. A ativação da RUBISCO carboxilase é feito pelo Mg++ e aumento do pH do estroma do cloroplasto. Durante a iluminação do cloroplasto, prótons são bombeados do estroma para o lúmen do tilocóide provocando um gradiente de prótons para a síntese de ATP nos cloroplastos necessários no ciclo de Calvin.Esse movimento de prótons é contrabalanceado pelo magnésio que faz o caminho inverso, isto é, do lúmem para o estroma para propiciar o equilíbrio eletrogênico e se tornar mais alcalino.
- Frutose 1,6 bisfosfatase: Esta enzima nos cloroplastos regula a síntese de amido e as partes que podem ser exportadas das células como as trioses fosfatos.
- Glutamina Sintetase (GS)Esta enzima cataliza a assimilação de amônio na reação com glutamato.Glutamato + NH4 → GS Glutamina. Portanto, tem papel fundamental na assimilação de nitrogênio.O nitrato (NO–3) é reduzido a nitrito (NO2) e posteriormente o nitrito é reduzido a NH4+ (amônio).O amônio (NH4+) através da enzima Glutamina Sintetase (GS) se liga ao glutamato produzindo glutamina.A glutamina é o metabolito chave para a produção de todos os outros aminoácidos que farão parte do metabolismo.Caso esta enzima (GS) não funcione a contento, por exemplo, pela deficiência de Mg++, há acumulo de amônio (NH4+) no cloroplasto. A acumulação de (NH4+) livre leva a uma desregulação do pH no lúmen do cloroplasto provocando o rompimento da membrana do cloroplasto, com sua potencial destruição levando a sintomas de necrose nas folhas.
OBS: O modelo de regulação na fixação de CO2 e a assimilação do nitrogênio são dependentes de Mg++.
Por outro lado, em plantas deficientes em magnésio, há acúmulo de carboidratos (amidos e açúcares) nas folhas das respectivas plantas, responsável pelo alto conteúdo de matéria seca nestas folhas, indicando que a fotossíntese é menos afetada que a degradação do amido nos cloroplastos afetando a produção de sacarose e sua respectiva translocação.
A acumulação de carboidratos em plantas deficientes em magnésio, se correlaciona com a diminuição dos teores de carboidratos nos tecidos drenos como vagens, por exemplo.
Este efeito também é notado no desenvolvimento das raízes, isto é, a diminuição de açúcares para as raízes que também são dreno, reduzindo seu crescimento. Este efeito deve-se provavelmente a problemas na enzima H+ ATPase que é a bomba de prótons para carregamento de sacarose na floema. Caso o magnésio seja suprido este efeito desaparece.
A acumulação de fotossintetatos nas folhas exerce uma regulação tipo “feedback” sobre a RUBISCO (carboxilase/oxidase) em favor da oxigenase, ativando a assimilação de O2 livre, consequentemente ativando o mecanismo de foto respiração que atrapalha a fotossíntese, isto é, a assimilação de CO2 que é feito pela porção carboxilase da enzima.
Assim devido à mudança no metabolismo do oxigênio em plantas deficientes em magnésio há acumulo de espécies reativas de O2 com os radicais super óxidos (O2–) e peróxido de hidrogeno (H2O2) o que pode provocar necroses nos tecidos e este efeito leve a acumulação de anti-oxidantes como o ascorbato e aumento da atividade da SOD (Super Oxido Dismutase) a fim de reduzir estas espécies tóxicas de O2.
Devido a este acúmulo o efeito de oxidação provocada pela luz podendo gerar necrose nos tecidos devido a foto–sensibilidade.
Potássio
O potássio é um cátion monovalente com raio hidratado de 0,33mm. Sua absorção é altamente seletiva e relacionada com a atividade metabólica da planta. O íon tem alta mobilidade nas plantas tanto em células individuais quanto em tecidos mais complexos. Pode ser transportado pelo xilema e pelo floema.
O potássio é o íon mais abundante no citoplasma juntamente com algum acompanhante como cloro ou nitrato o que lhe dá a característica de ser um regulador osmótico, portanto altamente correlacionado com os movimentos de água nas plantas.
O potássio não é metabolizado, portanto não faz parte da estrutura de moléculas ou enzimas. Portanto é altamente trocável dentro das células, não competindo em sítios de ligações como os cátions bivalentes (Mg++ e Ca++).
Suas principais funções estão descritas abaixo.
1. Por ser altamente solúvel e móvel e ter altas concentrações no citoplasma ele neutraliza outros solúveis como ácidos orgânicos, por exemplo, ou outros ânions inorgânicos solúveis como cloreto, assim estabilizando o pH citoplasmático entre 7,0 e 8,0, o que é o ideal para a maioria das reações metabólicas envolvendo outras moléculas orgânicas. Por exemplo, a do pH citoplasmático de 7,7 para 6,5 inibe completamente a atividade do nitrato redutase (enzima responsável pela assimilação do N).
2. A concentração do potássio no citoplasma e nos cloroplastos é sempre mantida muito alta em até 200 mM. No entanto a concentração nos vacúolos é baixa. Por outro lado, é extremamente alta nas células guarda dos estômatos podendo chegar a 500 mM. Assim, o potássio tem na regulação osmótica, uma de suas principais funções no metabolismo.
3. Ativação enzimática: Um grande número de enzimas é ativado pelo potássio.O potássio por ser monovalente induz mudanças na conformação molecular das enzimas aumentando a taxa de reação catalítica como a Vmax (Velocidade máxima de reação) e a constante Km que mede a afinidade da enzima no substrato.Em plantas deficientes em potássio ocorrem mudanças químicas nas enzimas levando a um acúmulo de carboidratos solúveis nos tecidos com consequente diminuição na síntese de amido e um acúmulo também de nitrogênio solúvel. Esta mudança no metabolismo de carboidratos deve estar relacionada ao alto requerimento de potássio (K+) para o funcionamento de algumas enzimas de regulação como, por exemplo, a piruvato quinase e a fosfopiruvato quinase.
- AMIDO SINTETASE. A atividade da enzima amido sintetase é altamente dependente da presença de cátions monovalentes principalmente o potássio sendo o mais afetivo. Esta enzima catalisa a transferência da glicose para as moléculas de amido. ADP – GLICOSE + AMIDO ⇄ ADP + GLICOSE – AMIDOOu seja, sem potássio não há síntese de amido, sem a síntese do amido não há produção de matéria seca, portanto a própria produtividade. Por outro lado, altas concentrações de K+ pode inibir a atividade da enzima.
- H+ ATPase – Enzima responsável pelas bombas de prótons. Esta enzima não somente é responsável por transportar o potássio do exterior para dentro da célula, mas faz do potássio o elemento essencial na regulação osmótica e na extensão celular.
- Hidrolases e Oxidases – Em planta com deficiência de potássio, os tecidos exibem alta atividade de enzimas proteolíticas como hidrolases e oxidases como a polifenol oxidase, em relação ás plantas normais. Em plantas deficientes há também acúmulo de “putrescina”. A enzima que catalisa a síntese de putrescina a partir de arginina é inibida sob altas concentrações de potássio, além de provocar o abaixamento do pH citoplasmático. Considerando o papel do potássio na manutenção do pH em níveis altos no citoplasma parece que o aumento da putrescina e reflexo da deficiência de potássio.
4. SÍNTESE DE PROTEÍNAS: O potássio, para a síntese de proteínas é mais solicitado do que para a própria ativação enzimática, ou seja, se requer muito mais potássio para a síntese das proteínas que simplesmente para sua ativação como antes se acreditava.O potássio esta envolvido na transcrição dos ribossomos quando da ligação do RNA transportador para a síntese das proteínas. Ou seja, o potássio esta envolvido na formação das ligações peptídicas na síntese de proteínas.Por exemplo, numa planta C3 a maioria das proteínas que se encontram nos cloroplastos é dirigida para a síntese da Ribulose, 1,5, bisfosfato carboxilase (RUBISCO), a principal enzima do Ciclo de Calvin envolvida na fotossíntese. A síntese dessa enzima é amplamente prejudicada na falta de K.Em plantas deficientes em K+, há acumulo de nitrogênio solúvel nas folhas, o que significa que este não esta sendo metabolizado em proteínas. É acumulado como aminas, aminoácidos e nitrato.Assim parece que o K+ é uma das principais chaves na ativação do próprio nitrato redutase, responsável pela assimilação do nitrato em compostos mais complexos, por outro lado, parece ainda provado que o K+ é a chave para a própria síntese dessa enzima.
5. FOTOSSÍNTESE: No processo de fotólise da água, quando há a liberação de H+ no estroma do cloroplasto o potássio é contra íon do processo, promovendo o fluxo de hidrogênio pela membrana do tilacóide e na estabilização do gradiente transmembrana, necessário para a produção de ATP, igualmente como ocorre na síntese de ATP nos mitocôndrias no processo de respiração.A presença de K+ também favorece a assimilação de CO2 na fotossíntese.Para a manutenção do pH alto no estroma, na presença de luz, há necessidade do influxo de K+ que vem citoplasma para dentro do cloroplasto e este influxo é mediado pelo balanço H+ / K+ para dentro do envelope do cloroplasto.Este balanço H+ / K+ é muito importante em condições de estresse hídrico. Durante a desidratação em condições de estresse hídrico, os cloroplastos perdem grande quantidade de K+ e a fotossíntese tende a diminuir. Esta diminuição na fotossíntese pode ser contra balanceada pela aplicação de K+ no citoplasma, ou seja, ainda fora do cloroplasto.Assim, mesmo em plantas intactas o efeito do estresse hídrico sobre a fotossíntese pode ser compensado pela aplicação de potássio. (K+).Desta forma o efeito do estresse hídrico sobre a fotossíntese e muito menos drásticos em plantas adequadamente supridas com potássio.
Esta alta demanda de potássio em plantas sob estresse hídrico ou salino se deve ao fato da regulação do pH no estroma, mantendo alta a presença do K+ no estroma nestas condições.
Portanto, em plantas deficientes, mesmo em condições normais, vários fatores relacionados à absorção de CO2 são afetados.
Deste modo, um aumento da presença de potássio nas folhas, aumenta a taxa de fotossintético e atividade da enzima RUBISCO carboxilase. Portanto, sob condições de boa nutrição potássica a assimilação de CO2 aumenta, consequentemente com a redução da foto respiração o que é comum em plantas deficientes.
Vale ainda ressaltar que além da atividade enzimática o potássio também regula a abertura e fechamento dos estômatos que em última análise também afeta a fotossíntese.
O aumento da disponibilidade de potássio reduz a respiração no escuro o que também afeta a taxa de fotossíntese liquida.
6. REGULAÇAO OSMÓTICA E EXTENSÃO CELULAR: A expansão celular é resultado da expansão do vacúolo, o qual ocupa de 80 a 90% do volume da célula.Para que haja a expansão celular há necessidade do aumento da extensibilidade da parede celular o qual esta envolvida com o crescimento ácido provocado pelo efluxo de H+ e redução do pH da parede, quebrando as pontes de cálcio nas pectinas e permitindo a expansão, e segundo a acumulação de solutos para aumentar o potencial osmótico, com consequente acumulação de potássio no vacúolo, o qual é requerido para estabilizar novamente o pH da parede para sua reconstrução.Portanto o efluxo de H+ produzido pela auxina para acidificar a parede é contrabalanceada pelo influxo de K+ para estabilizar o pH do citoplasma. Na deficiência de K+, a elongação induzida pela auxina (AIA) declina e cessa em poucas horas. Portanto em plantas deficientes a elongação celular é muito menor do que em plantas bem supridas em potássio.Paralelamente a extensão de caules provocada pelas giberilinas também é dependente do suprimento de potássio. Potássio e GA são sinergéticos. Isto é quando supridos em conjunto promovem maior elongação e crescimento.Assim, o potássio e outros íons como cloro, além de ácidos orgânicos são os principais ingredientes requisitados pelo vacúolo para a expansão celular. O potássio associado a açucares e ácidos orgânicos é o principal soluto requerido para a expansão celular. Dessa forma, o papel de toda as outras substâncias envolvidas na expansão celular dependem do status nutricional do potássio.
7. MOVIMENTO ESTOMÁTICO: Na maioria das espécies de plantas o potássio (K+) associado a um ânion (Cl–), por exemplo, tem a maior responsabilidade na mudança de conformação das células guarda dos estômatos durante os movimentos de abertura e fechamento.O aumento do K+ nas células guarda induz o aumento do potencial osmótico resultando em maior absorção de água pelas células e consequente aumento no turgor e assim provocando a abertura do estômato.O fechamento do estômato no escuro se processa de maneira inversa, isto é, o efluxo de potássio para fora a célula aumenta, ao contrário diminuindo a pressão de turgor com consequente desidratação pela diminuição de potencial osmótico e o estômato se fecha.
Resumo
Finalizando, gostaríamos de evidenciar que as principais funções fisiológicas das bases na vida das plantas estão aí destacadas. Não precisa ser um “expert” para entender o papel de cada base nutriente no estado nutricional das plantas, mas é necessário ter muito discernimento para valorizar a nutrição das plantas através do diagnóstico analítico, para só então tomar as decisões cabíveis na aplicação suplementar de nutriente, seja via foliar, para os micro, e/ou via radicular, para os macro.
Dessa forma, baseado nos informes acima a aplicação de nutrientes sem critérios de doses e ou quantidades, isto é, sem diagnóstico é um tanto temerário, podendo colocar as produções em risco não somente pela falta de nutrientes, mas também pelos seus excessos.
A revisão acima foi baseada em traduções deste autor do livro:
MARSCHNER, HORST. Mineral Nutrition of the Higher Plants. 2nd Edition. Academic Press. 1995.




Sou estudante de agronomia, estou no quinto ano do curso, a nutrição de plantas é o tema que pretendo me aprofundar. Obrigado pelas informações e por disponibilizar a literatura fonte usada.